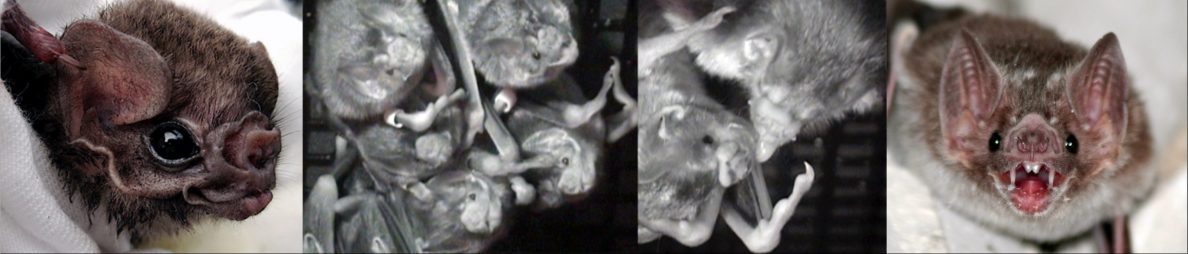
Spix’s disc-winged bat Thyroptera tricolor has suction cups (yes, suction cups) on its thumbs, and it uses these to cling to the smooth surface of young, furled Heliconia leaves.


I’ve wanted to see a disc-winged bat since I was about ten years old, and I finally got to see them this week while visiting Gloriana Chaverri, and Caroline and Michael Schöner at their field site in Costa Rica over the weekend.
Gloriana is a leading expert on social lives of Neotropical leaf-dwelling bats. This includes bats that chew leaves to make them into “tents” as well as disc-winged bats (Thyroptera). Over the last seven years, Gloriana has shown that Thyroptera live in small groups with multiple matrilines that often switch between their very temporary roosts, the furled leaves [1-3]. Both sons and daughters often remain at their natal site, and inbreeding is avoided because the bats mate with members of other populations. Each population includes several groups that switch among several roosts.
Despite moving daily between their tubular leaf roosts, the bats maintain very cohesive social networks. To coordinate their roost-switching, the bats use a system of contact calling [4-7]. Bats flying in search of their group make one contact call type called an “inquiry call” and groupmates within furled leaves make another call type in response. The calls have both individual and group-level signatures. The calls are also amplified by the funnel-like shape of the leaf roost.
Here’s a video of one flying into a leaf roost:
Most recently, Gloriana has been working with Caroline and Michael Schöner. The Schöners have pioneered work on an astonishing mutualism in another leaf-dwelling bat [8-10]. There is a pitcher plant in the paleotropics (Nepenthes) that has coevolved with a small insectivorous bat of the genus Kerivoula. The bat lives inside the pitcher and defecates into the fluid-filled “pitcher”. Most pitcher plants are carnivorous. They trap, drown, and digest insects inside the tiny pool of their pitcher, but this pitcher plant is modified to be a tiny little one-bat house. The level of the digestive fluid is lower to accommodate the bat’s roosting position. The plant lets the bat do the capturing, chewing, and pre-digesting of the insect prey. Some pitcher plants also attract shrews with a nectar reward and get them to poop into the pitcher [11]. For lack of a more polite term, the Schöners call the plants “coprophagous”.
In exchange for nutrients, the plant benefits the bat by providing it with a literal “roof over it’s head” and it’s distinctive shape is acoustically attractive to the bats. This is yet another example of bat-plant acoustic communication. There are also bat-pollinated flowers and leaves that reflect and enhance the echolocation calls of bats when they are ready to be pollinated [12-13].
What’s especially intriguing is that Kerivoula will also live in a different pitcher plant species that is not coevolved for the bats, and the bats can only fit inside the pitcher because it has been emptied. How? Something chews a small hole at the base of the pitcher to drain the liquid out. Could it be the bat? Nobody knows for sure. The Schöners suspect this is the case, but they have not yet caught the bat in the act. The bat might be a mutualist for the first species, but a parasite for the other. Or perhaps there’s a three-way interaction: the bat, the pitcher plant, and a third mystery organism that makes the hole, perhaps to drinks the pitcher plant liquid. It’s an unsolved mystery.


See also this blogpost by Merlin Tuttle and photos.
The Schöners are surprised by the physical similarities of Thyroptera and Kerivoula, although the personalities of the two bats are quite distinct. Kerivoula will eat larger insects and has the bite to match. Thyroptera it seems are a bit easier to work with. After your experiments are done, you can release Thyroptera into a furled leaf and they will just crawl inside and stay. As Caroline puts it, “Thyroptera is more well-behaved.”
The Schöners are tracking the bats’ movements among different leaves in the wild. Do the same bats consistently find new leaves and others follow? Or do all the bats do a search on their own? They are also looking at the bats use of spatial memory when relocating roosts. For this experiment, they work in a flight tent with an array of artificial leaves. The work is fascinating and I’m excited to see the results.
The Schöners take turns tromping around in the forest looking for bats and staying at the field station looking after their 9-month-old, Sophia, who loves fried platanos and has inherited the Schöners’ love of smiling. One similarity between Gloriana and the Schöners is their friendliness and propensity for hearty laughing. The atmosphere at the field station is one of joyful cheer, with everyone trying their best to make Sophie smile. It’s been a great place for a holiday.
Before coming to the Baru field station where I am now, I visited two caves with my wife Michelle, my labmate Nia Toshkova, and our guide: Gloriana’s graduate student Stanimira Deleva, who is studying the variation in use of caves by all different Costa Rican bats. The larger caves was two kilometers long and full of Pteronotus gymnonotus or Pternotus davyi. The smaller cave had Saccopteryx bilineata and Peropteryx kappleri. Nia and Stanimira are old friends, both cavers from Bulgaria. Michelle and I met Stanimira on a caving trip in Panama where I found a new potential site to work with vampire bats.
Here is a video of the Thyroptera being very cute:
References:
- Chaverri, G., & Kunz, T. H. (2011). All-offspring natal philopatry in a neotropical bat. Animal behaviour, 82(5), 1127-1133.
- Buchalski M, Chaverri G, and Vonhof M. 2014. When genes move farther than offspring: gene flow by male gamete dispersal in the highly philopatric bat species Thyroptera tricolor. Molecular Ecology 23:464-480.
- Chaverri G. 2010. Comparative social network analysis in a leaf-roosting bat. Behavioral Ecology and Sociobiology 64:1619-1630.
- Chaverri G, Gillam EH, and Vonhof MJ. 2010. Social calls used by a leaf-roosting bat to signal location. Biology Letters 6:441-444.
- Gillam EH, and Chaverri G. 2012. Strong individual signatures and weaker group signatures in contact calls of Spix’s disc-winged bat, Thyroptera tricolor. Animal Behaviour 83:269-276.
- Chaverri G, Gillam EH, and Kunz TH. 2012. A call-and-response system facilitates group cohesion among disc-winged bats. Behavioral Ecology 24:481-487.
- Chaverri G, and Gillam EH. 2013. Sound amplification by means of a horn-like roosting structure in Spix’s disc-winged bat. Proceedings of the Royal Society B 280:20132362.
- Grafe TU, Schöner CR, Kerth G, Junaidi A, and Schöner MG. 2011. A novel resource–service mutualism between bats and pitcher plants. Biology Letters 7:436-439.
- Schöner CR, Schöner MG, Kerth G, and Grafe TU. 2013. Supply determines demand: influence of partner quality and quantity on the interactions between bats and pitcher plants. Oecologia 173:191-202.
- Schöner MG, Schöner CR, Simon R, Grafe TU, Puechmaille SJ, Ji LL, and Kerth G. 2015. Bats are acoustically attracted to mutualistic carnivorous plants. Current Biology 25:1911-1916.
- Clarke, C. M., Bauer, U., Ch’ien, C. L., Tuen, A. A., Rembold, K., & Moran, J. A. (2009). Tree shrew lavatories: a novel nitrogen sequestration strategy in a tropical pitcher plant. Biology Letters, 5(5), 632-635.
- von Helversen, D., & von Helversen, O. (1999). Acoustic guide in bat-pollinated flower. Nature, 398(6730), 759-760.
- Simon, R., Holderied, M. W., Koch, C. U., & von Helversen, O. (2011). Floral acoustics: conspicuous echoes of a dish-shaped leaf attract bat pollinators. Science, 333(6042), 631-633.

Cool suction cups!
LikeLike