

“Direct reciprocity” occurs when individuals are more likely to help individuals that helped them previously, while “generalized reciprocity” (also called “upstream reciprocity“) occurs when individuals are more likely to help after they have been helped by any individual. So direct reciprocity is like “I’ll scratch your back if you scratch mine” and generalized reciprocity is like “pay it forward“. Although generalized reciprocity could be adaptive, this behavior could also be simply a nonadaptive byproduct of direct reciprocity. For example, a predisposition to caring about baby animals could be seen as a maladaptive byproduct of selection for caring about one’s own baby, and similarly, generalized reciprocity may result completely from selection for direct reciprocity. But others think generalized reciprocity may be very important in some animal societies because it poses fewer cognitive demands.
Unlike direct reciprocity, generalized reciprocity cannot stabilize cooperation all by itself, because there’s no way to prevent cheating in a well-mixed population.
I recently received an email asking how I distinguished between evidence of direct and generalized reciprocity in vampire bat food sharing. The email also pointed out that a minor error (when stating that I found no evidence of generalized reciprocity, I mistakenly referred to the supplementary materials even though supporting evidence was in the next few lines).
My reply was very long and spread across two emails, so I’m copying it below. I’ve added links to references where appropriate.
Hi Dr. Taborsky,
Thanks for your interest in our paper. I’m a big fan. I have been following your lab’s work on cooperative breeding in cichlids and generalized reciprocity in rats.
I’ll answer your second question first: “food received” in Fig. S4 refers to food received from a particular partner (not in general).
For your first point, I’m sorry for the confusion. The results supporting that statement are actually the sentences directly following and not in the supplement. I’m a bit embarrassed to say that I believe it’s a mistake. This text was moved to the supplementary materials at some point to ensure meeting the page limits, then they were moved back to the main text (so the reference to the supplement is mostly unnecessary). Here’s that text again:
Donation sizes could sometimes be compared both before and after the donor was fed within a round of trials. In these 28 cases, we failed to find a difference in presence of food sharing (paired t=0.98, df=27, p=0.34), total food donated (paired t=-1.3, df=27, p=0.20), or food donated per recipient (paired t=0.16, df=27, p=0.87). When the donor was fed on the previous day, we found no difference between the amount donated on that day compared to the donor’s average on other days (n=9 donors and 9 trials, paired t=-0.013, df=8, p=0.99).
However, there are some additional relevant things that you can get from the food sharing matrix in the supplement. For example, one can easily test if — among bats that both gave and received– overall food received predicts overall food given, which is consistent with generalized reciprocity. After log-transforming the data, there is a weak but NS trend between total help given and total help received by bats that did both (R-squared = 0.19, p=0.09). See correlation pictured below.
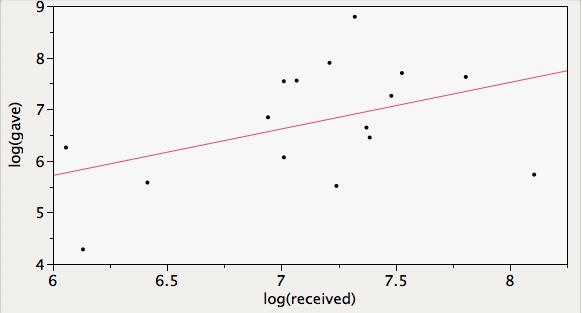
Some thoughts on evidence for generalized reciprocity in the study
I’m certainly interested in looking further at the role of generalized reciprocity in the vampire bats. Although I failed to find evidence that bats donated more often after they had recently received, my sample size was very small, so the effect of generalized reciprocity would have had to be substantial to be detected. Bats can certainly act as donors after being fasted and fed the previous night. My expectation was that the bats would be more likely to feed others after being a recipient in a trial. There’s actually two reasons for this. The first is the possibility of generalized reciprocity (bats might be more likely to share with others after having just received recently). The second reason is that the fasted bats always fed immediately after the food sharing trial (after all the other bats had already fed), and so of all potential donors for the next night, the previous day’s recipient had fed most recently, usually a large amount. However, I did not find any support for that prediction as I explained in the paper. I will expound on this in case it was unclear.
After a bat was fed by others, I found no subsequent change in the amount or probability of their subsequent food donations. There were 28 cases where a bat had opportunities to feed others before and after it served as a subject within a fasting round. When I compared food sharing before and after the donor was fasted, I found no significant difference for the total amount of food given by the donor, the amount of food given per recipient, or the probability of giving food during a trial. There were also 9 bats that donated to others the day after they were fed. In these 9 cases, the mean sharing on days immediately after being fed (327.6 s) did not exceed the mean sharing on other days (328.4 s). Of course, these sample sizes are also very small (and yes, I did just now see your editorial Ethology paper on sample sizes!). Basically, our data do not make a strong case for or against generalized reciprocity as a factor in food sharing, and of course we did not test the effect of anonymity and compare it with trials showing reciprocity (as you did with the rats). We did not find strong evidence of it.
Our data do not actually provide conclusive evidence for or against direct reciprocity either in my opinion. We are in the process of organizing more controlled tests in a lab setting to address this. A major advantage of working with vampire bats is that no training is required for the bats to share food, but the huge difficulty has been getting them into our lab. I have spent the majority of my PhD trying to do this, with limited success. I just brought my first group of 7 bats here 10 days ago.
Some thoughts on generalized reciprocity as an evolutionary explanation for food sharing
I think my results disprove the harassment idea and the idea that non-kin sharing only occurred due to a lack of kin presence. However, distinguishing between direct reciprocity and kin discrimination based on familiarity is far more difficult. Unlike these alternatives, I don’t consider generalized reciprocity to be a stand-alone alternative explanation, but rather as a probable supplementary mechanism, i.e. an additional source of information for decision-making as outlined by your “hierarchical information hypothesis” (Rutte and Taborsky 2008).
I don’t think generalized reciprocity can by itself explain food sharing in vampire bats because of the instability of roosting group membership. The vampire bat social network involves a lot of roost switching. As your lab showed previously, generalized reciprocity works well in stable groups of 3 where the actions and outcomes of generalized reciprocity are the same as direct reciprocity 50% of the time. (i.e. if you return anonymous help in a group of 3, there’s a 50% chance you are directly reciprocating).
However, generalized reciprocity strategies are self-promoting via passive positive assortment (or spatial selection, population viscosity, limited dispersal, or whatever you want to call it), so generalized reciprocity starts to breakdown with more social mixing. You showed this mathematically, but I think its also logically intuitive. Hence, indiscriminate helping towards with groupmates (whether interpreted as kin selection or equivalently multi-level selection) and indiscriminate helping based on past anonymous help both make the same assumptions and predictions when considered as a stand-alone explanation for cooperation. The assumption is that enforcement is not required due to population viscosity. And the prediction is that the animals should not be basing helping acts on individual discrimination of any kind.
Conclusion
Generalized reciprocity would be neither necessary nor sufficient to stabilize cooperative food sharing in vampire bats. In contrast, the other alternative mechanisms I discussed (e.g. enforcement via direct reciprocity or kin selection via limited dispersal or via nepotism) could in theory explain food sharing by themselves. That’s why I looked for observational evidence of generalized reciprocity, but dismissed it as an alternative explanation.
I think multiple factors and their interactions are at work, and I suspect generalized reciprocity probably helps promote cooperation alongside the other more key drivers. Indeed, before I started in his lab, I emailed Jerry Wikinson and noted that in his original 1984 small captive trials, there were 2 cases where starved bats were not fed by reciprocators and in both cases, the donors were the bats that had just been fed (by another bat) in the last trial. So generalized reciprocity has been on my mind since the beginning, but I have not find any real evidence for it yet.
Best
Gerry

You are awesome Gerry. Keep up the good work!
LikeLike